Steven Miller1, Victor Amadi2, Aschalew Z. Bekele3, Ulrike Zieger4, Harry Hariharan2and Diana Stone2
1Graduate Studies Program, School of Veterinary Medicine, St. George’s University, Grenada, West Indies
2Pathobiology Academic Program, School of Veterinary Medicine, St. George’s University, Grenada, West Indies
3Department of Veterinary Population Medicine, College of Veterinary Medicine, University of Minnesota, St. Paul, MN, United States
4Anatomy, Physiology and Pharmacology Academic Program, School of Veterinary Medicine, St. George’s University, Grenada, West Indies
Volume 2014 (2014),
Article ID 676408,
International Journal of Veterinary Medicine: Research & Reports,
6 pages,
DOI: 10.5171/2014.676408
Received date: 18 December 2013; Accepted date: 17 March 2014; Published date: 23 April 2014
Academic Editor: Elzbieta K Jagusztyn-Krynicka
Cite this Article as:
Steven Miller, Victor Amadi, Aschalew Z. Bekele, Ulrike Zieger, Harry Hariharan and Diana Stone (2014), "Identification of Human and Poultry Campylobacter Sequence Types in Small Indian Mongooses (Herpestesauropunctatus) in Grenada, West Indies," International Journal of Veterinary Medicine: Research & Reports, Vol. 2014 (2014), Article ID 676408, DOI: 10.5171/2014.676408
Campylobacter is one of the leading causes of bacterial diarrhea in humans. Poultry and other domestic food animals are the primary reservoir hosts for Campylobacter species that infect humans. The mongoose in Grenada is one of most abundant mammals on the island and lives in close proximity to humans and livestock. Thus it was of interest to determine if the mongooses in Grenada harbor human pathogenic, antibiotic-resistant Campylobacterstrains. In the present study, two of 82 (2.4%) mongooses were positive for Campylobacter jejuni based on rectal swab cultures. From these two positive samples a total of five isolates were obtained and genotyped. Three of the five isolates were obtained in pure culture and tested for antibiotic susceptibility. The two isolates which could not be purified from contaminating Gram-positive cocci were excluded from susceptibility tests. All three pure isolates tested were susceptible to ciprofloxacin, erythromycin, tetracycline, chloramphenicol, ampicillin, gentamicin and metronidazole as determined by E-test. All isolates tested were susceptible to ciprofloxacin, erythromycin, tetracycline, chloramphenicol, ampicillin, gentamicin and metronidazole as determined by E-test. MLST results revealed two clonal complexes (CCs) and three sequence types (STs) among the five isolates: clonal complex ST-354 containing sequence type ST-6196 and clonal complex ST-464 containing sequence types ST-6729 and ST-464. Both CCs have been identified in humans and poultry worldwide and it is possible that mongoose acquisition of these strains may contribute to human exposure.
Campylobacter is one of most common causative agents of bacterial food and waterborne illness in humans (Sheppard et al., 2009) and is responsible for an estimated 2.4 million cases of human illness per year in the US alone (Thomas et al., 1999; Center for Disease Control and Prevention, (CDC), National Center for Emerging Zoonotic and Infectious Diseases, 2010). Campylobacter is a gram negative, non-spore forming, rod-shaped bacterium and the genus is comprised of at least 17 species. Most Campylobacter species are microaerophilic, and most pathogenic species require thermophilic conditions for growth. The most common species affecting humans is C. jejuni. However, among several other species, C. coli,C. lariand C. ureolyticus have also been implicated in human infection (Thomas et al., 1999; Allos, 2001; Koziel et al., 2012). Poultry are considered a major source of human infection; however, other livestock such as cattle, sheep and pigs have also been identified as potential reservoirs forCampylobacter species (Aarestrup et al., 1997) and unpasteurized ruminant milk is a recognized source of human infection (Allos, 2001).
Inter and intra species diversity within the Campylobacter genus makes identification difficult at the species level and below when using non-molecular techniques (Sheppard et al., 2012). Multi locus sequence typing (MLST) is one of the most effective methods for genotyping of Campylobacter spp. It is a method of genotyping which uses seven housekeeping genes, approximately 400 bp each. The use of seven loci for identification allows for high discriminatory power when genotyping (Colles et al., 2003). Until recently, the use of MLST was considered time consuming and expensive. However new advances have made this technology more affordable and time efficient (Sheppard et al., 2012).
Grenada is a small volcanic island in the south Caribbean approximately 344 km² in size (Central Intelligence Agency, 2013). Mongooses were introduced to Grenada in the late 1800’s, mostly for rodent pest control and have been introduced in various countries worldwide over the last 200 years. In Grenada, the mongoose has become one of the most abundant mammals on the island with numbers estimated near 200,000 (Horst et al., 2001). Mongooses live in close proximity to humans and livestock and have a preference for preying on poultry. In addition, mongooses in Grenada serve as reservoirs for several human pathogens including rabies and Salmonella (Everard and Everard, 1988; Miller et al., 2013).
The purpose of the present study was to determine: (1) the prevalence of Campylobacter jejuni/coli/larigroup in mongooses in Grenada, (2) the Campylobacter species and sequence types present among these isolates from mongooses and (3) the antibiotic resistance profiles of these sequence types to 7 antimicrobial drugs. In Grenada, antibiotic resistance has been seen in C. jejuni strains isolated from several animals including chickens, pigs, sheep and goats and it was of interest to determine if mongooses harbor Campylobacter strains with similar resistance profiles.
Materials and Methods
Mongoose Sampling
Mongooses were live trapped from all six parishes in Grenada as part of a rabies surveillance program (IACUC 10003-R). A total of 82 mongooses were sampled from September 2012 to March 2013 during both the wet and dry season. Animals were anesthetized within two hours of collection via intra-muscular injection with ketamine (10mg/kg) (Ketamine hydrochloride, Rotexmedica®, Trittau, Germany) and xylazine (0.2 mg/kg) (AnaSed®, Decatur, IL) and then euthanized via cardiac injection of potassium chloride (1-2 mmol/kg) (Fisher Scientific, Fair Lawn, NJ).
Sample Collection and Campylobacter Isolation
Fecal sample for Campylobacter isolation were taken using sterile Cary Blair transport swabs (BBL, Becton, Dickinson and Company, Sparks, MD, USA) which were inserted into the rectum promptly after euthanasia and plated within two hours of collection on Campylobacter blood-free media (CBF) (Oxoid LTD) using quadrant streaking techniques. Cefoperazone (Bio-Merieux Inc.) and amphotericin B supplements (Bio-Merieux Inc.) were added to the CBF medium to inhibit growth of other enteric bacteria. The plates were incubated under microaerophili cconditions usingCampylobacter gas generating kit (Campy-Pak, Oxoid LTD, Basingstoke, Hampshire, England) gas packs at 42° C for 48 hours. After incubation, colonies exhibiting typical Campylobacter morphology (flat, grey, mucoid) were selected for Gram staining (Fisher Scientific Co., Kalamazoo, MI). After morphological confirmation, Campylobacter colonies were sub-cultured on CBF plates to obtain pure cultures. Catalase, oxidase and hippurate tests (Remel, Lennexa, KS, USA) were performed, along with latex agglutination for Campylobacter jejuni, Campylobacter coliand Campylobacter lari (JCL, Integrated Diagnostics, Baltimore, MD, USA) groups. Sensitivity tests against cephalothin and nalidixic acid were conducted as previously described (Hariharan et al., 2009). Isolates showing sensitivity to nalidixic acid, resistance to cephalothin and positive results for hippurateand JCL agglutination tests were identified as C. jejuni. Pure cultures were placed in 10% sterile milk in cryovials (Thermo Fisher Scientific Inc.) and stored at -80° C until genotyping.
Multi-locus Sequence Typing (MLST)
Stock cultures of the isolates were grown on blood agar plates for 24 h at 42°C under microaerophilic conditions. Total DNA was extracted using DNA blood and tissue purification kits (DNeasy Blood & Tissue Kits, Qiagen Inc., Valencia, CA, USA) according to manufacturer’s instructions and samples were shipped to the College of Veterinary Medicine, University of Minnesota, St. Paul, MN for MLST determination. PCR amplification of seven genes was carried out as described previously (Dingle et al., 2001, 2002; Schouls et al., 2003) using guidelines and PCR primer sequences obtained from the Campylobacter PubMLST database (http://pubmlst.org/campylobacter/mlst-info/Cjejuni/primers.html). Briefly, PCR was performed using commercial ready-to-use master mixes in a master cycler (Mastercycler (Eppendorf AG), HotStarTaq® Master Mix kit, Qiagen Inc., Valencia, CA, USA). The reaction mixture contained 10 µM of each primer, 25 µL of PCR master mix and 2 µL of template DNA and water to make total volume of 50 µL. The cycling program consisted of an initial denaturation at 95°C for 15 min, followed by 35 cycles of 94°C for 30 sec, respective annealing temperature for each primer for 1 min and extension at 72°C for 3 min with a final extension at 72°C for 10 min. To confirm the presence of amplicon of the expected size, the PCR amplification products were run on 1.2% agarose gel. Purified PCR products were quantified and sequenced (ACGT, Inc., Wheeling, IL, USA) using the same primers used for the PCR amplification. The obtained sequences were analyzed using sequence analysis software (Sequencherver 10.1, Gene Codes Corporation, MI, USA). Sequence data were submitted to the Campylobacter PubMLST database for allele assignments.
Antibiotic Susceptibility Testing
Antibiotic susceptibility testing using the E-test (AB Biodisk, Solna, Sweden) and interpretation of results were performed using established methods (Hariharan et al., 2009). The E-test was used for overall resistance testing since no internationally agreed upon susceptibility criteria for Campylobacter species currently exist (Varela et al., 2008). The following antibioticswere used for susceptibility testing: ampicillin, chloramphenicol, ciprofloxacin, erythromycin, gentamicin, tetracycline and metronidazole (Difco/BD).
Results
Prevalence and Antibiotic Susceptibility
Only two of 82 (2.4%) mongooses were positive for Campylobacter in the present study and all isolates were C. jejuni. From these two positive samples a total of five Campylobacter isolates were obtained. Up to three expectedCampylobacter colonies were sub-cultured from each positive mongoose to determine if a single mongoose could carry multiple Campylobacter species and/or genetic clones. Only three of the five isolates were tested for antibiotic susceptibility because the two remaining isolates exhibited Gram-positive contaminants along with Campylobactersas determined by Gram stain and could not be isolated in pure culture after five streak plate attempts on CBF selective medium with antibiotics. All isolates tested were susceptible to ampicillin, chloramphenicol, ciprofloxacin, erythromycin, gentamicin, tetracycline and metronidazole.
Molecular Typing
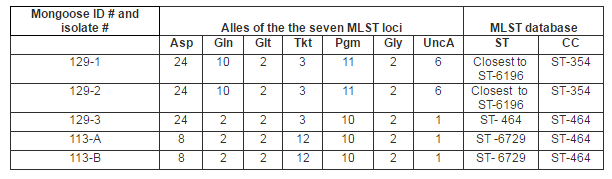
Campylobacter MLST procedures only amplify target gene loci of Campylobacter species. Thus, all five isolates from the two mongooses, including those from the non-pure cultures, were submitted for MLST typing. MLST results revealed two clonal complexes (CCs) and three sequence types (STs) among the five isolates: CC ST-354 containing ST-6196 and CC ST-464 containing ST-6729 and ST-464 (Table 1). One of the two mongooses was positive for both CC ST-354 (ST-6196, two isolates) and CC ST-464 (ST-464, one isolate), while both isolates of the other mongoose were CC ST-464 (ST-6729).
Table 1. MLST Profiles of Campylobacter Isolates from Two Mongooses
Discussion
Both of the CCs identified in the present study have been reported in poultry in Grenada (Stone et al., 2013) but not in goats, sheep or pigs in Grenada (Stone, personal communication, 2013). Small ruminants are not part of the mongoose’s diet and the overall prevalence of Campylobacter was low among sheep and goats (4.5% and 3.2% respectively) in Grenada suggesting environmental contamination with these small ruminant STs is also low. Worldwide, CC ST-354 is isolated predominantly from humans and chickens and has been reported in multiple countries (PubMLST, 2013). ST-6196, the only ST found within CC ST-354 in mongooses in Grenada has previously only been reported in Japan (PubMLST, 2013). However the source of the isolate was not reported. The second complex, CC ST-464, contained the only ST found in mongooses that was also reported in poultry in Grenada, ST-464 (Stone et al., 2013). This likely reflects the fact that poultry are a common prey animal of mongooses in Grenada. Currently 208 PubMLST records exist for ST-464, most of which have been isolated from humans and poultry. There is only one PubMLST record for ST-6729 from CC ST-464 which was isolated from human stool in Thailand. Based on current PubMLST data, CC ST-354 and CC ST-464 are most commonly isolated from humans and poultry worldwide.
To date, antibiotic resistance has been reported in C. jejuni strains isolated from chickens, pigs, sheep and goats in Grenada (Hariharan et al., 2009; Ganchingco et al., 2012; Roopnarine et al., 2013; Stone, pers. communication, 2013). In particular, ST-464 isolated from poultry in Grenada exhibited resistance to tetracycline (Stone et al., 2013). However the same ST isolated from mongoose in the present study showed no resistance to tetracycline. Again, only three of the five isolates in the present study were isolated in pure culture and tested for antibiotic susceptibility. If mongooses primarily acquire Campylobacter strains from poultry and if antibiotic resistance increases among these strains, then increased antibiotic resistance should also be seen among poultry-associated strains isolated from mongooses in the future. Although our findings are novel, further studies with large number of isolates may be required to elucidate the antimicrobial resistance patterns of Campylobacters from mongooses.
In conclusion, although overall prevalence was low, the identification of human pathogenic Campylobacter strains in mongooses in Grenada suggests that wildlife species known to prey on Campylobacter reservoir hosts may need to be considered in the overall ecology and epidemiology of Campylobacter. Both of the clonal complexes identified in the present study have previously been reported in poultry in Grenada, and in humans and poultry in many countries worldwide. Because poultry are a common source of human pathogenic Campylobacter strains, it is possible that mongoose acquisition of these strains may contribute to human exposure. Most research on Campylobacter in animals has focused on domestic livestock species. Future studies investigating the prevalence, sequence types and antibiotic resistance profiles of Campylobacter in wildlife, particularly wildlife species that live in close proximity to humans and livestock, will contribute to our understanding of Campylobacter.
Acknowledgements
This study had the approval of the St. George’s University Institutional Animal Care and Use Committee (IACUC 10003-R) and was conducted as part of an active rabies surveillance program. We acknowledge Dr. Alfred Chikweto, Dr. Keshaw Tiwari and Mr. Ray Samuels of St. George’s University, School of Veterinary Medicine, Grenada, and property owners of the various collection sites in Grenada for their assistance in this research.
Miller, S., Amadi, V., Stone, D., Johnson, R., Hariharan, H. & Zieger, U. (2014). ‘Prevalence and Antimicrobial Susceptibility of Salmonella Spp. in Small Indian Mongooses (Herpestes Auropunctatus) in Grenada, West Indies,’Comparative Immunology, Microbiology and Infectious Diseases, submitted Feb 2014.
PubMLST (2013). ‘Campylobacter Multi Locus Sequence Typing Home Page,’ [Online] Available from: http://pubmlst.org/ campylobacter. [Accessed 5th Oct. 2013]